
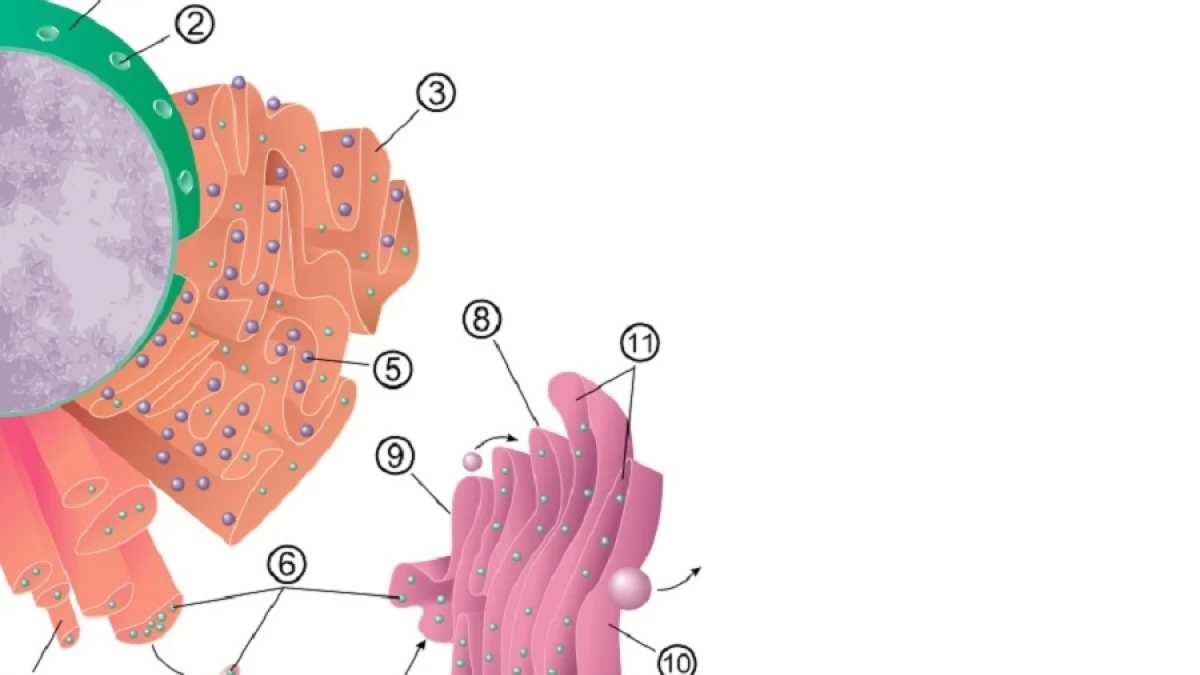

El retículo endoplasmático es un órgano celular formado por membranas interconenctadas. Estas membranas se continúan con las del centro de la célula, el núcleo celular.
Existen dos tipos de retículo endoplasmático: uno, denominado rugoso, cuyas membranas forman cisternas aplanadas y con ribosomas asociados, y el otro llamado liso, el cual se organiza con membranas formando túbulos sin ribosomas.
En este artículo vamos a hablar sobre el retículo endoplasmático rugoso, cuales son sus partes y sus funciones.
- Artículo relacionado: "Tipos de células principales del cuerpo humano
¿Qué es el retículo endoplasmático rugoso?
Este orgánulo, además de retículo endoplasmático rugoso, recibe otros nombres: retículo endoplasmático granular, ergastoplasma o retículo endoplásmico rugoso. Este orgánulo solo se puede encontrar en células eucariontes.
Estructuralmente, se caracteriza por estar formado por una serie de canales, sacos aplanados y cisternas, los cuales se encuentran distribuidos por el medio de la célula, el citoplasma.
En estos sacos aplanados se introducen cadenas hechas por varios péptidos, con los cuales se formarán proteínas complejas. Estas mismas proteínas viajan a otras partes de la célula, como lo es el aparato de Golgi y el retículo endoplasmático liso.
Alrededor de los sacos que forman este orgánulo se encuentran numerosos ribosomas asociados a ellos. Estas estructuras son unas vesículas que pueden contener proteínas y otras sustancias. Estos ribosomas son los que le dan una apariencia rugosa al ser observado por el microscopio.
Esta estructura tiene por función principal la de sintetizar proteínas, las cuales son destinadas a distintas partes de la célula para desarrollar múltiples funciones, además de controlar su calidad estructural y funcional.
Funciones
Estas son las principales funciones del retículo endoplasmático rugoso.
1. Síntesis de proteínas
El retículo endoplasmático rugoso tiene una función que es de vital importancia para la supervivencia del organismo: sintetizar proteínas.
Estas proteínas pueden desempear múltiples funciones, ya sean estructurales, formando parte de otros orgánulos, actuando como hormonas, enzimas o sustancias transportadoras. Así pues, el lugar de destino de estas proteínas puede ser el propio interior de la célula en donde han sido sintetizadas, formando la capa celular o yendo hacia el exterior de esa célula.
La mayoría de las proteínas que forman parte de los orgánulos de la célula tienen su origen en los ribosomas del retículo endoplasmático. Esta síntesis alcanza su fase final en el interior del retículo endoplasmático rugoso.
El proceso da comienzo cuando se unen ácido ribonucleico mensajero (ARNm) a una unidad ribosómica pequeña y, luego, a una de grande. Así se da inicio al proceso llamado traducción.
Lo primero que se traduce es la secuencia de nucleótidos, la cual sintetizará una cadena de unos 70 aminoácidos. Esta cadena recibe el nombre de péptido señal. Una molécula llamada SRP (sequence recognition particule) se encarga de reconocer a este péptido señal, enlenteciendo el proceso de traducción.
La estructura formada por las dos subunidades ribosómicas, el ARNm, el péptido señal y la SRP viaja por el citosol hasta llegar a la pared del retículo endoplasmático rugoso.
Mediante una proteína especial, llamada translocador, se forma un canal en la membrana por el que pasa la parte peptídica de la estructura formada. El péptido señal se une al translocador, el resto de la cadena peptídica se va traduciendo e introduciendo al interior del retículo.
Una enzima, llamada peptidasa, rompe el péptido señal del resto de la cadena de aminoácidos, quedando esta cadena libre en el interior del orgánulo.
Una vez la síntesis ha sido completada, la cadena de aminoácidos va adquiriendo una estructura tridimensional, propia de una proteína completa, y se va plegando.
- Quizás te interese: "Los 20 tipos de proteínas y sus funciones en el organismo"
2. Control de calidad
El retículo endoplasmático rugoso lleva a cabo una función fundamental para el buen funcionamiento orgánico. Este orgánulo ejerce un importante papel a la hora de detectar proteínas con defectos o que puedan no resultar útiles para el organismo.
El proceso empieza cuando se detecta una proteína que ha sido mal plegada a la hora de ser sintetizada. Las enzimas encargadas de esta fase del proceso son el grupo de las glucosiltransferasas.
La glucosiltransferasa le añade una glucosa a la proteína defectuosa, concretamente en su cadena de oligosacáridos. El objetivo de esto es que una chaperona, concretamente la calnexina, reconozca la glucosa de esta proteína y la detecte como proteína mal formada, así la regresará a su lugar de origen para que sea bien plegada.
Este proceso ocurre varias veces. En caso de que no se dé la corrección por esta vía, se pasa a la siguiente fase.
La proteína es dirigida a una parte llamada proteosoma, en donde será degradada. En este lugar trabajan múltiples tipos de enzimas las cuales desintegran la proteína defectuosa en aminoácidos que se pueden reciclar para formar una nueva proteína bien plegada.
Esta función de control de calidad y detección de aquello sintetizado que no es útil o que incluso puede resultar ser tóxico para la célula cumple una función higiénica muy importante.
Así pues, la célula puede encargarse de asegurarse de que las proteínas bien formadas llegan al punto de maduración en el que son funcionales, mientras las que no son desechadas o recicladas.
Variedades de ergastoplasma
En función de la célula en la que se encuentre, este orgánulo presenta unas características estructurales diferenciadas, y también cabe la posibilidad de que reciba otro nombre.
En las células secretoras, el retículo endoplasmático rugoso se manifiesta en forma de numerosas cadenas o sacos dispuestas en paralelo y poco separadas entre sí, lo suficientemente como para que se puedan formar las vesículas con las que se sintetizan sustancias.
En el sistema nervioso, este orgánulo recibe el nombre de cuerpos de Nissl, apareciendo en forma de cisternas muy separadas con muchos ribosomas libres por el citosol. Algunas neuronas, pese a tener este orgánulo, apenas sintetizan proteínas.
Referencias bibliográficas:
- English, A. R., Zurek, N., Voeltz, G. K. (2009). Peripheral ER structure and function. Current opinion in cell biology, 21, :506-602.
- Daleke D. L . (2007). Phospholipid Flippases. The journal of biological chemistry. 282, 821-825.
- Nixon-Abell J, Obara, C. J., Weig V. A., Li D., Legant W. R., Xu C. S., Pasolli H. A., Harvey K., Hess H. F. , Betzig E., Blackstone C., Lippincott-Schwartz3 J.( 2016). Increased spatiotemporal resolution reveals highly dynamic dense tubular matrices in the peripheral ER. Science. 354, 3928-2.













